玉兰小鬼伞Coprinellus magnoliae
N.I. de Silva, Lumyong & K.D. Hyde 2021
基本信息 | - 通用中文名Chinese name:玉兰小鬼伞
- 别称&俗称Othernames:木兰小鬼伞
- 拉丁学名Scientific name:Coprinellus magnoliae N.I. de Silva, Lumyong & K.D. Hyde 2021
- 科family:小脆柄菇科Psathyrellaceae
- 属Genus:小鬼伞属Coprinellus
- 分布Distribution:中国、泰国
- 有分布的省区Provinces:广东省,海南省,贵州省,广西壮族自治区,云南省(文献记载)
查看具体分布情况 - 生态Ecology:仲春或夏季群生或簇生于苔藓、土壤、阔叶树腐木上,或作为植物内生菌。
- 营养类型Nutrition Type:腐生Saprotrophic: 分解死亡生物体或有机物获取养分
- 置信指标Confidence:★★★
|
介绍部分
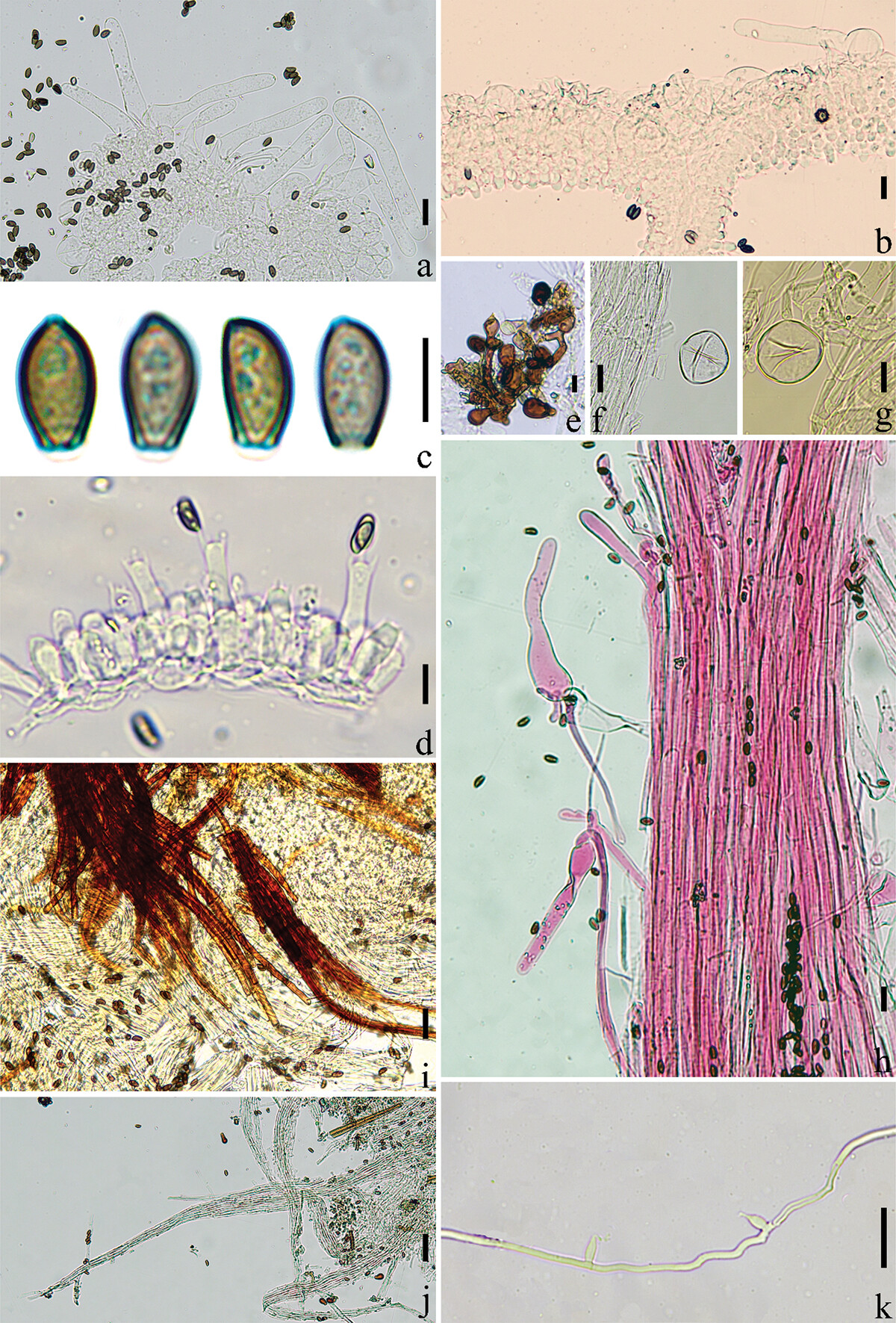
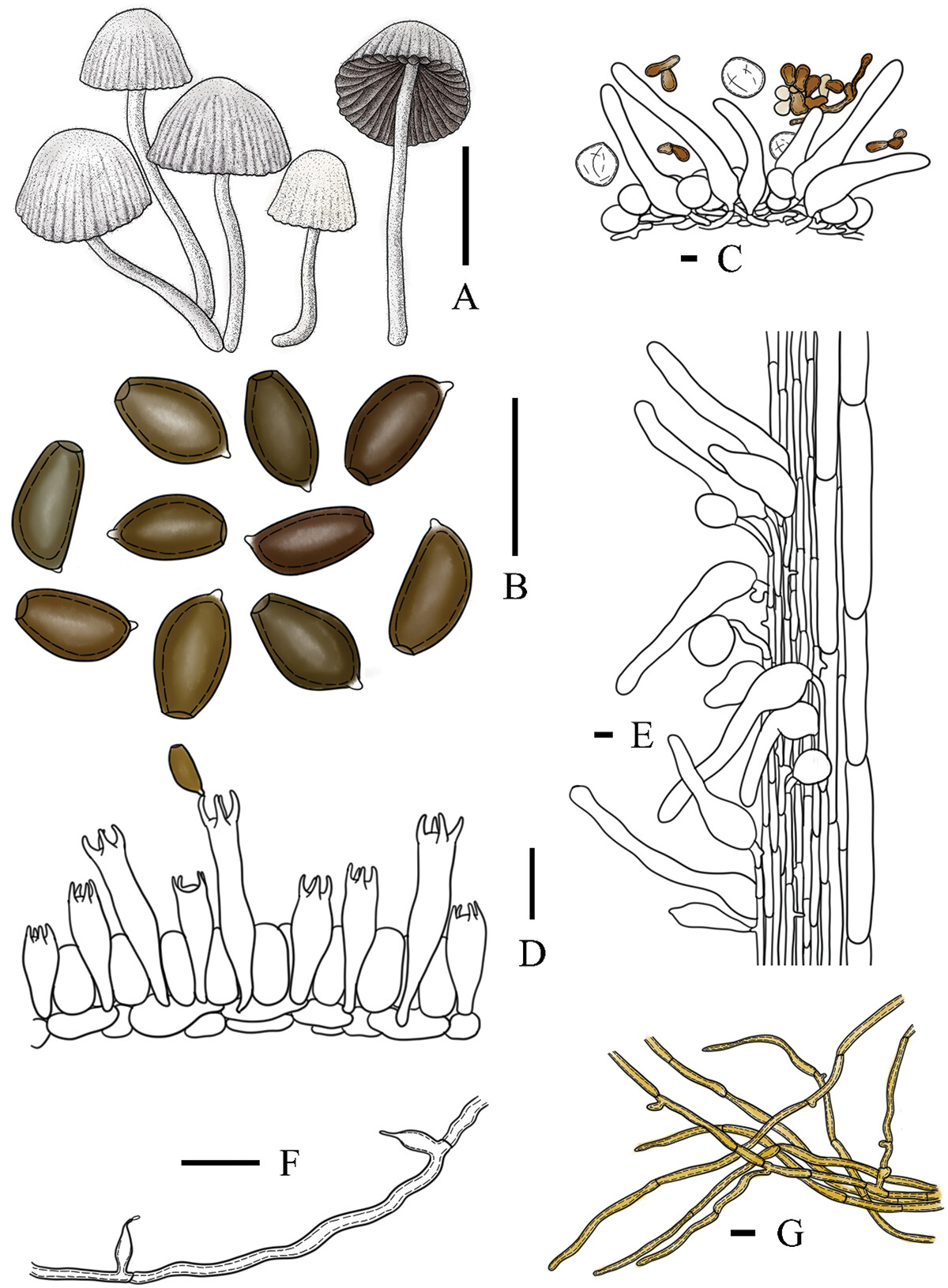
简介:玉兰小鬼伞是分布于亚洲亚热带和热带地区的一种小鬼伞,是白小鬼伞复合群的成员。类似于其他白小鬼伞复合群的成员,其担子果小型,常大量簇生;菌盖有沟纹,灰色调;菌褶成熟后褐色调;菌柄细长而脆;存在盖生囊状体。它的鉴别性特征是:菌盖成熟后常常带有一定的紫色调(呈紫灰色),菌盖中部没有黄褐色调;柄生囊状体相比其他物种更长(通常120 − 170μm,最短82μm),在菌柄的下部也可以找到;主要分布在广东、贵州、云南、海南等省份。
发现历程
该物种最早是在Ni de Silva等(2021)对中国和泰国地区的植物内生菌(endophytic fungi)研究中发现的。他们从泰国玉兰
Magnolia garrettii中分理出一种内生真菌的菌丝体,测序结果表面它是小鬼伞属的一种,因此命名为玉兰小鬼伞。
2024年,L. Y. Zhu等报道了该物种在我国华南地区的广泛分布,也首次描述了其子实体的宏观特征。
相似物种
绒柄小鬼伞
C. velutipes和玉兰小鬼伞近缘,但其菌柄表皮有长、分支的末端细胞。
微小鬼伞
C. parcus的菌柄下部有特征性的近球形或囊状末端细胞,有别于玉兰小鬼伞。
物种介绍引证的参考文献
(上文介绍部分中所引用的参考文献)
- Species clarification of fairy inkcap (“Coprinellus disseminatus”) in China, 2024. Liyang Zhu & Tolgor Bau. Mycology https://doi.org/10.1080/21501203.2024.2309901
- Morpho-molecular taxonomic studies reveal a high number of endophytic fungi from Magnolia candolli and M. garrettii in China and Thailand, 2021. Ni de Silva et al.. Mycosphere 12(1): 163–237 https://doi.org/10.5943/mycosphere/12/1/3
涉及到该物种的相关文献
(能证明该种在我国有分布的文献)
- Species clarification of fairy inkcap (“Coprinellus disseminatus”) in China, 2024. Liyang Zhu & Tolgor Bau. Mycology https://doi.org/10.1080/21501203.2024.2309901
- Morpho-molecular taxonomic studies reveal a high number of endophytic fungi from Magnolia candolli and M. garrettii in China and Thailand, 2021. Ni de Silva et al.. Mycosphere 12(1): 163–237 https://doi.org/10.5943/mycosphere/12/1/3
序列信息
如何下载序列?
| 分子类型 |
标本号 |
Version No. |
序列 |
文献 |
来源地 |
| DNA | HMJAU 46307 | OR436399 | ITS | ↗ | 中国 广东省 Shaoguan City, Chebaling National Nature Reserve on moss layer |
| DNA | HMJAU 46307 | OR436381 | LSU | ↗ | 中国 广东省 Shaoguan City, Chebaling National Nature Reserve on moss layer |
| DNA | HMJAU 49305 | OR436400 | ITS | ↗ | 中国 海南省 Baisha County, Wuzhishan National Nature Reserve on rotten wood in broad-leaved forest |
| DNA | HMJAU 49305 | OR436382 | LSU | ↗ | 中国 海南省 Baisha County, Wuzhishan National Nature Reserve on rotten wood in broad-leaved forest |
| DNA | HMJAU 67122 | OR436401 | ITS | ↗ | 中国 广西壮族自治区 Baise City, Leye Courty, Yachang Orchid National Nature Reserve, Niuping Protection Station clayed soil in broad-leaved forest |
| DNA | HMJAU 67122 | OR436383 | LSU | ↗ | 中国 广西壮族自治区 Baise City, Leye Courty, Yachang Orchid National Nature Reserve, Niuping Protection Station clayed soil in broad-leaved forest |
| DNA | MFLU 20–0605[主模式] | MW244022 | ITS | ↗ | 泰国 Chiang Mai Province Mueang Chiang Mai District healthy leaves of Magnolia garrettii (Magnoliaceae) |
物种描述
以下描述摘自相关参考文献,仅供参考。可能存在文献过时、描述错误或者描述片面等情况
文字识别时可能有纰漏,描述若与参考文献原文有出入以原文为准
外文描述的参考翻译系网站管理者的个人翻译,仅供参考,具体以原文为准
描述1 描述来源:Species clarification of fairy inkcap (“Coprinellus disseminatus”) in China类型:完整的描述(摘录自专著、论文等)
描述原文:
Coprinellus magnoliae N.I de Silva, Lumyong & K.D. Hyde, Figures 14–16
Description: Basidiomata tiny-sized. Pileus 0.4 − 0.7 cm, first ellipsoid, semisphere when mature, without a rounded umbo; (sub)white (3A1) when young, become grey-white (5B1) or light purple-brown (7B2) to purple-grey (9D2) at age, without brown hue at the centre; the radial grooves extended to the top of the cap after maturation, with a clear line at the folding part which is slightly darker than other parts; pubescent. Context very thin, only present at the centre part, white (1A2) to cream (1A3). Lamellae narrow adnate, L = 16 − 19, I = 1, 0.1 − 0.2 cm in wide, first white (1A2) to light grey-white (9B1), finally dark brown-grey (7E3), usually deliquescent at age especially in moist environment. Stipe 2.8 − 4.0 × 0.1 cm, white (1A1) to cream (1A3), hollow, crispy, pubescent, without a volva-like margin.
Basidiospores [80, 4, 4] (6.2) 7.4 − 7.6 (8.4) × (3.9) 4.3 − 5.4 (4.7) × (3.4) 4.1 − 4.3 (4.6) μm, Q1 = 1.55 − 1.91, Q2 = 1.62 − 2.04, long ovoid in frontal view, amygdaliform with acute apex in side view, obconical at base and truncate at apex, dark red-brown (6E5) in water and brown-grey (6E3) in 5% KOH solution; germ pore central to slightly eccentric, 0.8 − 1.8 μm in wide. Basidia trimorphologic, 13 − 31 × 4 − 7 μm, short clavate to clavate, sometimes bulging in middle part, 4- or 2- spored, sterigmata 3 − 5 μm in length; each basidium surrounded with 4 − 6 pseudoparaphyses. Cheilocystidia and pleurocystidia absent. Hymenophoral trama regular, trama hyphae 3 − 9 μm in wide. Pileipellis spherocystoderm at ridge part and groove part, consisting of subglobose, ellipsoid or utriform cells, 22 − 53 × 14 − 52 μm, hyaline, thin-walled; pileocystidia clavate, lageniform to slender lageniform, 38 − 183 × 11 − 29 μm, with 10 − 17 μm wide, with subcapitate apex, hyaline, thin-walled. Caulopellis hyphae 3 − 6 μm wide, hyaline, sometimes branched and diverticulate, thin-walled; hyphae of stipe trama 11 − 26 μm wide, hyaline, thin-walled to somewhat thick-walled. Caulocystidia clavate to narrow lageniform, 82 − 170 × 14 − 67 μm, with 6 − 16 μm wide, could be found until the lower part of the stipes. Elements of the veil have two types: 1) chains of ellipsoid to subglobose elements, 19 − 38 × 12 − 25 μm, thick-walled, cream (1A3) to dark brown (6E8); 2) free subglobose to globose elements, 18 − 45 × 13 − 41 μm, thin-walled, hyaline. Hyphae of ozonium parallel dense arrangement, 2 − 5 μm, slightly thick-walled to thick-walled, cream (1A3) to yellow-brown (5C6), darker in 5% KOH solution. Clamp connection and pseudoclamps are absent. The conidiophores are short and unbranched, constricted at the septa with swollen bases, hyaline, thick-walled, and around 2 μm in wide, the apical cell of each conidiophore could produce one subulate phialide, 8 − 9 × 3 − 4 μm, with a puncta apex. Conidium unseen.
Habitat: In groups or caespitose soil, moss layer, or rotten wood in broad-leaved trees. Usually occurs in the middle of spring to summer.
Distribution: Southern and eastern Asia (China, Thailand).
Additional specimens examined: China: Guangdong Province: Shaoguan City, Chebaling National Nature Reserve, on rotten wood in broad-leaved forest, 7 May 2017, Tolgor Bau and Mei Huang, HMJAU46308, HMJAU46311; same location, on moss layer, HMJAU46307 (ITS: OR436399; LSU: OR436381); Hainan Province: Baisha County, Wuzhishan National Nature Reserve, on rotten wood in broad-leaved forest, Tolgor Bau and Qin Na, 6 June 2018, HMJAU49305 (ITS: OR436400; LSU: OR436382); Guizhou Province: Qiandongnan Mao and Dong Autonomous Prefecture, Leigong Mountain National Forest Park, dead branches of broad-leaved tree, Jun-Qing Yan; Guangxi: Baise City, Leye Courty, Yachang Orchid National Nature Reserve, Niuping Protection Station, clayed soil in broad-leaved forest, Xiao-Liang Liu, Guang-Fu Mou and Li-Yang Zhu, 25 June 2021, HMJAU67122 (ITS: OR436401; LSU: OR436383); Yunnan Province: Kunming City, on rotten woods, October, 2017, Hai-Ying Bau, HMJAU49302; Kunming City, Kunming Botanical Garden, Fuligong Greenhouse, humus layer of Alocasia macrorrhizos, Tolgor Bau and Li-Yang Zhu, NaN Invalid Date NaN, HMJAU67135.
Note: This species was initially identified as an asexual endophytic fungus isolated from healthy leaves of Magnolia garrettii in Thailand and here we present the first description of its sexual stage. The main recognition feature of this species is the purple-grey pileus when mature, with the centre of the pileus lacking a brownish-yellow hue. Additionally, the caulocystidia of this species is relatively longer compared to other species in this section, typically measuring 120 − 170 μm in length, and could also be found even at the lower part of stipes. C. velutipes is close to C. magnoliae, however, it has long-branched terminal cells of caulopellis (105 − 190 μm in length) at the lower part of the stipes as well as clamp connection and pseudoclamp. The lower part of the stipes of C. parcus has distinct subglobose or utriform terminal cells that could be distinguished from C. magnoliae.
参考翻译:
描述:担子果微小。菌盖直径0.4~0.7厘米,初期椭圆形,成熟时呈半球形,无圆形脐突;幼时近白色(3A1),随成熟变为灰白色(5B1)或浅紫褐色(7B2)至紫灰色(9D2),菌盖中央无棕黄色调;成熟后放射状沟纹延伸至菌盖顶部,菌盖折叠处具清晰条纹,颜色略深于其他部分;被短绒毛。菌肉极薄,仅存在于中央部分,白色(1A2)至奶油色(1A3)。菌褶窄附生,菌褶数目L=16~19,小菌褶数目I=1,宽0.1~0.2厘米,初期白色(1A2)至浅灰白色(9B1),最终呈暗棕灰色(7E3),成熟后尤其在潮湿环境中通常液化。菌柄2.8~4.0 × 0.1厘米,白色(1A1)至奶油色(1A3),中空,脆,被短绒毛,无菌托状边缘。
担孢子 [80/4/4] (6.2)7.4 ~ 7.6(8.4)×(3.9)4.3 ~ 5.4(4.7)×(3.4)4.1 ~ 4.3(4.6)微米,Q1 = 1.55 ~ 1.91,Q2 = 1.62 ~ 2.04,正面观呈长卵形,侧面观呈杏仁形且顶端尖锐,基部倒圆锥形,顶端平截,在水中呈暗红棕色(6E5),在5% KOH溶液中呈棕灰色(6E3);芽孔位于中心至略偏心,宽0.8 ~ 1.8微米。担子三型(#指担子的长度集中聚集在3个区间内——网站作者注#),13 ~ 31 × 4 ~ 7微米,短棒状至棒状,有时中部膨大,4孢梗或2孢梗,担子小梗长3 ~ 5微米;每个担子周围环绕4 ~ 6个周细胞。褶缘囊状体和侧生囊状体无。菌髓规则型,菌髓菌丝宽3 ~ 9微米。菌盖表皮在脊部和沟部均为spherocystoderm型,由近球形、椭圆形或坛形细胞组成,22 ~ 53 × 14 ~ 52微米,透明,薄壁;盖生囊状体棒状,瓶状至细长瓶状,38 ~ 183 × 11 ~ 29微米,顶部宽10 ~ 17微米,具近头状顶端,透明,薄壁。柄生皮菌丝宽3 ~ 6微米,透明,有时分枝且具侧枝,薄壁;菌柄菌髓菌丝宽11 ~ 26微米,透明,薄壁至稍厚壁。柄生囊状体棒状至窄瓶状,82 ~ 170 × 14 ~ 67微米,顶部宽6 ~ 16微米,在菌柄下部亦可发现。菌幕组织有两种类型:1) 椭圆形至近球形细胞组成的链状结构,19 ~ 38 × 12 ~ 25微米,厚壁,奶油色(1A3)至暗棕色(6E8);2) 游离的近球形至球形细胞,18 ~ 45 × 13 ~ 41微米,薄壁,透明。菌索菌丝平行密集排列,宽2 ~ 5微米,稍厚壁至厚壁,奶油色(1A3)至黄棕色(5C6),在5% KOH溶液中颜色变深。无锁状联合和假锁状联合。分生孢子梗短且不分枝,在分隔处缢缩,基部膨大,透明,厚壁,宽约2微米,每个分生孢子梗的顶端细胞可产生一个钻形瓶梗(分生孢子器),8 ~ 9 × 3 ~ 4微米,顶端具孔。未观察到分生孢子。
生境:群生或簇生于阔叶林下的土壤、苔藓层或腐木上。通常出现于春末至夏季。
分布:南亚和东亚(中国、泰国)。
注:该物种最初被鉴定为从泰国M. garrettii健康叶片中分离出的一种无性内生真菌,本文首次描述了其有性阶段。该物种的主要识别特征是成熟时菌盖呈紫灰色,且菌盖中央无棕黄色调。此外,与本组其他物种相比,该物种的柄生囊状体相对较长,通常长120 ~ 170微米,并且在菌柄下部亦可发现。绒柄小鬼伞与木兰小鬼伞相近,但后者菌柄下部具长分枝的柄生皮顶层细胞(长105 ~ 190微米),并且具有锁状联合和假锁状联合。微小鬼伞菌柄下部具有明显的近球形或坛形顶层细胞,可与木兰库恩菇区分。
库中图片 3 张
标签:[无]
上次编辑:2025-11-29
物种编号:1056
Revised:true
本页向外链接的参考文献:总计 7 条链接(含重复)
重定向至本页的页面:总计 0 条链接
©菌物志Mycopedia
回到首页 搜索物种 目录
©菌物志Mycopedia,版权声明
如需引用本文可参考下列格式:
蔡佳铭.2025. 玉兰小鬼伞Coprinellus magnoliae. 菌物志Mycopedia. http://www.mycopedia.top/result.php?details=Coprinellus magnoliae
Jia-Ming Cai.2025. Coprinellus magnoliae [Chinese]. Mycopedia. http://www.mycopedia.top/result.php?details=Coprinellus magnoliae